Advocacy &Community &Diagnosis &Education &Elimination &Epidemiology &India &IRS &ITNs &Malaria &Plasmodium/Parasite &Procurement Supply Management &Research &Strategy &Surveys &Vaccine &Vector Control &World Malaria Day Bill Brieger | 24 Apr 2025
The Johns Hopkins Bloomberg School of Public Health Dean’s Lecture on Malaria
Professor Jane Carlton, PhD, Director of Johns Hopkins Malaria Research Institute was introduced by Dean Ellen MacKenzie to give a Dean’s Lecture entitled “Malaria: History, current status, and the promise of ‘omics and AI.”

Prof. Carlton first gave an overview of JHMRI, which was founded in 2001. She stepped into director role in 2023. She started with the encouraging premise that AI and ‘omics can supercharge our research and pointed out the power of comparative genomics on understanding parasites and disease. Her goal is to translate discovery into real world impact through collaborations.
The talk started with a brief malaria history. Malaria was described as an ancient disease and remains one of top infectious diseases worldwide. There were 2.63 million cases and more than half a million deaths in 83 endemic countries in 2023. Today 44 countries are malaria-free.
Up until now, Prof. Carlton noted, the malaria map has been shrinking. There was a precipitous decline in malaria in India from 23 million cases to 2 million. The disease is a true humanitarian issue with a large impact on people living in resource-limited settings where housing is basic and offers no protection from mosquitoes.

The JHMRI is supported by Bloomberg philanthropies, and from that base faculty research examines, among others, better methods for controlling mosquitoes (see slide), new diagnostic tests and therapeutics, and the next generation of vaccines. Key assets to support research include mosquito insectaries and a malaria parasite core. The insectaries produce 60,000 mosquitoes per week, and with these it is possible to complete the life cycle in the laboratory.
Continuing education is another important function of JHMRI which has three conferences per year including the upcoming World Malaria Day 2025 symposium. One can also learn from the Malaria Minute podcast. The upcoming “Vector Encounter” provides sharing and learning for researchers.

JHMRI studies malaria at field sites in Africa and Asia where country collaborators are partners. Emphasis is on local capacity building in countries like Zambia, Ethiopia, Kenya, and Uganda. Researchers and the national malaria control programs in these countries work hand-in-hand.
JHMRI is involved in developing the next generation of vaccines. One approach if a human monoclonal antibody that prevents malaria infections. Another develops a vaccine that transcends’ malaria parasite strains with structure guided mimicry of an essential P. falciparum receptor-ligand complex enhances cross neutralizing antibodies. A third example asks “How many parasites does it take to cause malaria?” and assesses infection likelihood through mosquito parasite burden.
To understand the theme of her talk, Prof. Carlton reviewed the promise of ‘omics and AI in the context of her work at a center of excellence in India. Pioneering work using Malaria camps in hard-to-reach villages in Odisha, India. The main activities mobilized villagers to gather for mass screening, treatment, education, and intensified vector control. From there, Indoor Residual Spraying was planned and insecticide treated bednets were distributed. Other maternal and child health activities were incorporated. After three rounds/visits in the remote villages a great drop of malaria cases was seen. WHO lauded the camps.

In addition to lessons about the importance of surveillance, mixed strategies, and community mobilization for controlling malaria, the team learned about the growing challenge of reduced effectiveness of Rapid Diagnostic Tests. The problem arose because tests were dependent on a protein that was no longer being expressed due to Pfhrp2 gene deletions, leading to false negative test results. The team was encouraged to identify more proteins to find a more stable and central one to use in testing. Through machine learning, this work is ongoing but promising.
While we are on the verge of several research and programmatic breakthroughs, Prof. Carlton reminded the audience that we are in calamitous times. She recalled that the United States has been the top donor government to malaria efforts through Presidents Malaria Initiative and Global Fund to Fight AIDS, Tuberculosis, and Malaria. PMI was founded in 2005, and has contributed to a decline in malaria death rates of close to 50%.

With suspension of funding, an estimated increase of 12.5-17.9 million malaria cases and 71,000-166,000 malaria deaths are expected this year. Already there are serious impacts on the supply chain for major malaria commodities as estimated by the Roll Back Malaria Partnership as seen on their RBM dashboard and supply chain gap estimates where six endemic countries have less than a 3-month supply of RDTs and eleven have less than a 6-month supply.
Prof Carlton ended by saying, “I think the hope is in science, right? The hope is in research. There are definitely new initiatives, new tools which are coming to the forefront, some of which I mentioned, and several of which we’re developing here at the malaria Research Institute. I do know the World Health Organization has got together with other countries to provide additional funding and support for those countries who have lost support through PMI.”
Case Management &Children &Efficacy &Household &Plasmodium/Parasite &Severe Malaria &Transfusion Bill Brieger | 21 Oct 2020
Malaria News Today 2020-10-21: perspectives on falciparum, transfusion issues, drug effectiveness
Ironically, blood transfusion helps with severe malaria, but can be dangerous in mild cases. And with severe malaria delay in treatment is a major risk. Malaria parasites can be surprisingly diverse, even in one home. Health system performance, drug quality and patient adherence are key factors in the effectiveness of anti-malarials. Read more on each article or abstract in the links provided.
The impact of delayed treatment of uncomplicated P. falciparum malaria on progression to severe malaria
 Mousa A et al. conducted a systematic review and a pooled multicentre individual-patient meta-analysis in PLoS Medicine. Despite the reported association of delay in receiving treatment for uncomplicated malaria (UM) with an increased risk of developing severe malaria (SM), access to treatment remains low in most high-burden areas. Researchers performed a pooled individual-participant meta-analysis with the aim to ascertain the correlation between treatment delay and presenting with SM using Ovid MEDLINE and Embas.
Mousa A et al. conducted a systematic review and a pooled multicentre individual-patient meta-analysis in PLoS Medicine. Despite the reported association of delay in receiving treatment for uncomplicated malaria (UM) with an increased risk of developing severe malaria (SM), access to treatment remains low in most high-burden areas. Researchers performed a pooled individual-participant meta-analysis with the aim to ascertain the correlation between treatment delay and presenting with SM using Ovid MEDLINE and Embas.
Findings revealed significantly higher risk of severe disease in children and adults who had longer delays from symptom onset to treatment-seeking; this relationship was noted to be the strongest for progression to severe malarial anaemia. Per estimates, nearly half of the severe anaemia cases in both children and adults could be averted if they presented within the first 24 hours of symptom onset.
Malaria parasites in Nigeria are genetically diverse: a danger but also a useful tool
Segun Isaac Oyedeji notes that his team’s research has already confirmed that in malaria-endemic countries such as Nigeria, infected individuals carry P. falciparum parasites that are genetically complex or diverse. What we didn’t know was how diverse the parasites are in the micro environment, such as within households and among children of the same family.
Oyedeji thought that knowing the population structure within households could help us understand more about the pattern and development of the disease. It could also inform development of appropriate guidelines and control measures. He found that even in the micro environment, P. falciparum parasites exhibit high genetic diversity. This finding was similar to results from larger communities in malaria endemic regions and has the same important implications. The implication is that a one-size fits all intervention or approach against the parasites may not be effective. There were children living under the same roof and infected by parasites that were genetically different.
New evidence to guide the practice of blood transfusions in children with severe malaria
The Barcelona Institute for Global Health (ISGlobal) described a new study that shows that transfusions could help increase survival, even at higher haemoglobin levels than those currently recommended. The results show that blood transfusion increased the survival of patients with severe disease.
In cases with complications, such as impaired consciousness or elevated lactate in blood, transfusion improved survival even in children whose levels of haemoglobin were higher the recommended threshold of 60g /l. For example, among patients with impaired consciousness, the authors observed improved survival upon transfusion with haemoglobin levels as high as 105 g / l. However, in the case of mild cases, transfusion was associated with an increase in mortality.
Global estimation of anti-malarial drug effectiveness for the treatment of uncomplicated Plasmodium falciparum malaria 1991–2019
 Giulia Rathmes and colleagues note that anti-malarial drugs play a critical role in reducing malaria morbidity and mortality, but their role is mediated by their effectiveness. Effectiveness is defined as the probability that an anti-malarial drug will successfully treat an individual infected with malaria parasites under routine health care delivery system. Anti-malarial drug effectiveness (AmE) is influenced by drug resistance, drug quality, health system quality, and patient adherence to drug use; its influence on malaria burden varies through space and time. This study used data from 232 efficacy trials.
Giulia Rathmes and colleagues note that anti-malarial drugs play a critical role in reducing malaria morbidity and mortality, but their role is mediated by their effectiveness. Effectiveness is defined as the probability that an anti-malarial drug will successfully treat an individual infected with malaria parasites under routine health care delivery system. Anti-malarial drug effectiveness (AmE) is influenced by drug resistance, drug quality, health system quality, and patient adherence to drug use; its influence on malaria burden varies through space and time. This study used data from 232 efficacy trials.
The global effectiveness of artemisinin-based drugs was 67.4% (IQR: 33.3–75.8), 70.1% (43.6–76.0) and 71.8% (46.9–76.4) for the 1991–2000, 2006–2010, and 2016–2019 periods, respectively. The use of artemisinin-based combination therapy (ACT) with a competent partner drug and having multiple ACT as first-line treatment choice sustained high levels of effectiveness. High levels of access to healthcare, human resource capacity, education, and proximity to cities were associated with increased effectiveness. Effectiveness of non-artemisinin-based drugs was much lower than that of artemisinin-based.
This study provides evidence that health system performance, drug quality and patient adherence influence the effectiveness of anti-malarials used in treating uncomplicated falciparum malaria. These results provide guidance to countries’ treatment practices and are critical inputs for malaria prevalence and incidence models used to estimate national level malaria burden.
Asymptomatic &Children &coinfection &IPTp &ITNs &Malaria in Pregnancy &Plasmodium/Parasite &Reproductive Health &Schools &Seasonal Malaria Chemoprevention Bill Brieger | 15 Oct 2020
Malaria News Today 2020-10-15
Recent publications in Malaria Journal, The Lancet and eLife tackle several challenges to saving lives and malaria elimination. Problems include low access to bednets for children in Ethiopia, high prevalence of asymptomatic malaria in Ghanaian adults, risk of co-infection with other infectious diseases, and gaps in current interventions to prevent malaria in pregnancy and children. On the hopeful side, new targets for drug therapy are being identified. Read more on each by following the links below.
Long-lasting insecticide-treated bed net ownership, utilization and associated factors among school-age children in Southern Ethiopia
Zerihun Zerdo and colleagues examined net use among children in malaria-prone areas of

Dara Mallo and Uba Debretsehay districts because malaria is one of the major causes of morbidity and mortality among school-age children (SAC) in sub-Saharan Africa. This study was part of a baseline assessment in a cluster-randomized controlled trial.
The ownership of at least one LLIN by households of school-aged children (SAC) was about 19.3% (95% CI 17.7–21.0%) but only 10.3% % (95% CI 7.7–13.7%) of these households had adequate access of bed nets to the household members. Ownership of bed net was lower than universal coverage of at least one bed net for two individuals. It is important to monitor replacement needs and educate mothers with low education level with their SAC on the benefit of consistent utilization of bed nets.
Prevalence of and risk factors for Plasmodium spp. co-infection with hepatitis B virus: a systematic review and meta-analysis
Kotepui and Kotepui observed that Plasmodium spp. and hepatitis B virus (HBV) are among the most common infectious diseases in underdeveloped countries. Therefore they examined co-infection in people living in endemic areas of both diseases. The PubMed, Web of Science, and Scopus databases were searched. Observational cross-sectional studies and retrospective studies assessing the prevalence of Plasmodium species and HBV co-infection were examined. and found 22 studies to include in a systematic review and meta-analysis. Overall, the pooled prevalence estimate of Plasmodium spp. and HBV co-infection was 6% (95% CI 4–7%, Cochran’s Q statistic?<?0.001, I2: 95.8%).
No difference in age or gender and risk of Plasmodium spp. and HBV co-infection group was found. The present study revealed the prevalence of Plasmodium spp. and HBV co-infection, which will help in understanding co-infection and designing treatment strategies. Future studies assessing the interaction between Plasmodium spp. and HBV are recommended.
High prevalence of asymptomatic malaria infections in adults, Ashanti Region, Ghana, 2018
Melina Heinemann and co-researchers noted that Ghana is among the high-burden countries for malaria infections and recently reported a notable increase in malaria cases. While asymptomatic parasitaemia is increasingly recognized as a hurdle for malaria elimination, studies on asymptomatic malaria are scarce, and usually focus on children and on non-falciparum species. Therefore asymptomatic adult residents from five villages in the Ashanti Region, Ghana, were screened for Plasmodium species by rapid diagnostic test (RDT) and polymerase chain reaction (PCR) during the rainy season. Samples tested positive were subtyped using species-specific real-time PCR.
 Molecular prevalence of asymptomatic Plasmodium infection was 284/391 (73%); only 126 (32%) infections were detected by RDT. While 266 (68%) participants were infected with Plasmodium falciparum, 33 (8%) were infected with Plasmodium malariae and 34 (9%) with P. ovale. The sub-species P. ovale curtisi and P. ovale wallikeri were identified to similar proportions. Non-falciparum infections usually presented as mixed infections with P. falciparum.
Molecular prevalence of asymptomatic Plasmodium infection was 284/391 (73%); only 126 (32%) infections were detected by RDT. While 266 (68%) participants were infected with Plasmodium falciparum, 33 (8%) were infected with Plasmodium malariae and 34 (9%) with P. ovale. The sub-species P. ovale curtisi and P. ovale wallikeri were identified to similar proportions. Non-falciparum infections usually presented as mixed infections with P. falciparum.
Most adult residents in the Ghanaian forest zone are asymptomatic Plasmodium carriers. The high Plasmodium prevalence not detected by RDT in adults highlights that malaria eradication efforts must target all members of the population. Beneath Plasmodium falciparum, screening and treatment must also include infections with P. malariae, P. o. curtisi and P. o. wallikeri.
Scientists shed new light on mechanisms of malaria parasite motility
eLife reports a new insight on the molecular mechanisms that allow malaria parasites to move and spread disease within their hosts has just been published. The first X-ray structures of the molecular complex that allows malaria parasites to spread disease highlight a novel target for antimalarial treatments.
The movement and infectivity of the parasite Plasmodium falciparum, and ultimately its ability to spread malaria among humans, rely on a large molecular complex called the glideosome. The new findings provide a blueprint for the design of future antimalarial treatments that target both the glideosome motor and the elements that regulate it.
New Lancet Series: Malaria in early life
Malaria infections are harmful to both the pregnant mother and the developing fetus. Malaria is associated with a 3–4 times increased risk of miscarriage and a substantially increased risk of stillbirth, and it disproportionately affects children younger than 5 years. Falciparum malaria is responsible for more than 200 000 child deaths per year in Africa and vivax malaria causes excess mortality in children in Asia and Oceania. In a duet of papers, we review 1) the deleterious effects of malaria in pregnancy on the developing fetus and 2) the current strategies for prevention and treatment of malaria in children.
Paper 1 is “Deleterious effects of malaria in pregnancy on the developing fetus: a review on prevention and treatment with antimalarial drugs” by Makoto Saito, Valérie Briand, Aung Myat Min, and Rose McGready. The authors are concerned that one in ten maternal deaths in malaria endemic countries may result from Plasmodium falciparum infection, that malaria is associated with a 3–4 times increased risk of miscarriage and a substantially increased risk of stillbirth. While current treatment and prevention strategies reduce, but do not eliminate, malaria’s damaging effects on pregnancy outcomes. They conclude that there is a need for alternative strategies to prevent malaria in pregnancy.
Paper 2 is “Treatment and prevention of malaria in children” by Elizabeth A Ashley and Jeanne Rini Poespoprodjo. They examine the following interventions: Triple antimalarial combination therapies, the RTS,S/AS01 vaccine, seasonal malaria chemoprevention and preventing relapse in Plasmodium vivax infection with primaquine.
Advocacy &coronavirus &Fever &HIV &IPTp &Journalists/Media &Malaria in Pregnancy &Plasmodium/Parasite &Private Sector &Tuberculosis Bill Brieger | 06 Oct 2020
Malaria News Today 2020-10-06: malaria in pregnancy, parasites surviving fever and private sector support
Today’s news featured a media briefing by the RBM Partnership and AMMREN on the challenges of protecting pregnant women from malaria. Additional news expands on these challenges. Research looks at how malaria parasites withstand the heat of a patient’s fever. Finally examples are presented of collaboration between international organizations and the private sector for malaria and disease control. Follow the links to gain more information.
Speed Up IPTp Scale-Up: a media briefing on maternal health
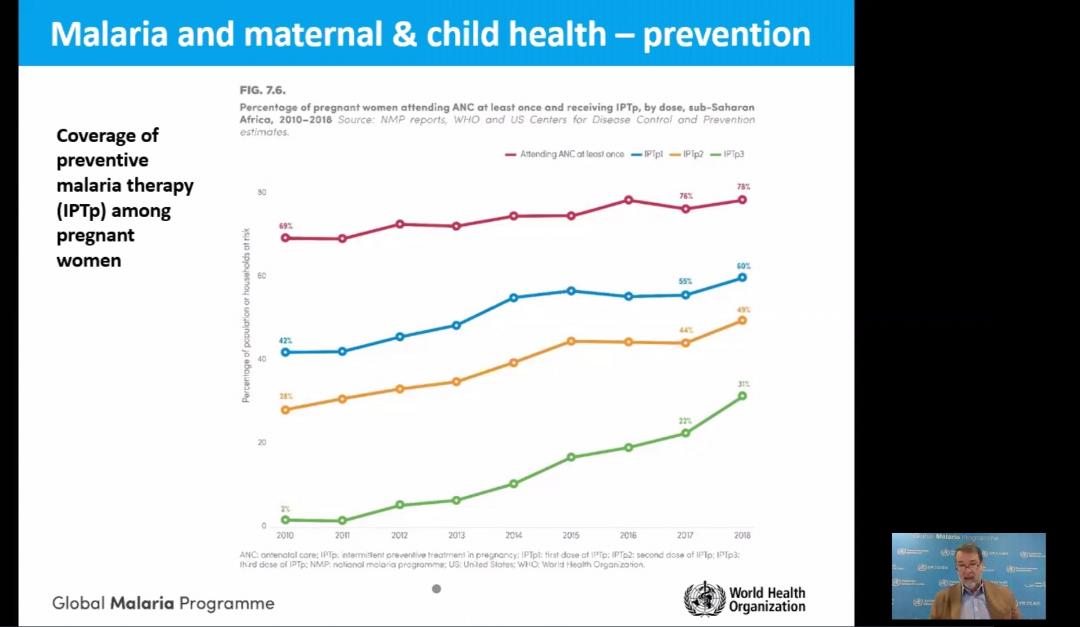 A panel discussion and media briefing on new approaches and lessons learned formed part of an online global call to end malaria in pregnancy with intermittent preventive treatment. The briefing on Tuesday, 6 October 2020, was sponsored by the Roll Back Malaria Partnership and AMMREN. The RBM website features background on the call to action.
A panel discussion and media briefing on new approaches and lessons learned formed part of an online global call to end malaria in pregnancy with intermittent preventive treatment. The briefing on Tuesday, 6 October 2020, was sponsored by the Roll Back Malaria Partnership and AMMREN. The RBM website features background on the call to action.
The panel discussion included Dr. Aminata Cisse ep. Traore, Sous Directrice de la Santé de la Reproduction/Direction Générale de la Santé et de l’Hygiène Publique, Ministére de la Santé et de l’Hygiène Publique, Mali; Dr. Anshu Banerjee, Director Department of Maternal, Newborn, Child, Adolescent Health & Ageing, World Health Organization (WHO), and Dr. Pedro Alonso, Director Global Malaria Programme, WHO. The discussion was moderated by Mildred Komey, Malaria in Pregnancy Focal Person, National Malaria Control Program, Ghans Health Service.
 The discussion covered the importance of launching this call now, what we’ve learned over the last few years, and examples of innovative strategies from Mali. There was a Q&A session with journalists after the presentations.
The discussion covered the importance of launching this call now, what we’ve learned over the last few years, and examples of innovative strategies from Mali. There was a Q&A session with journalists after the presentations.
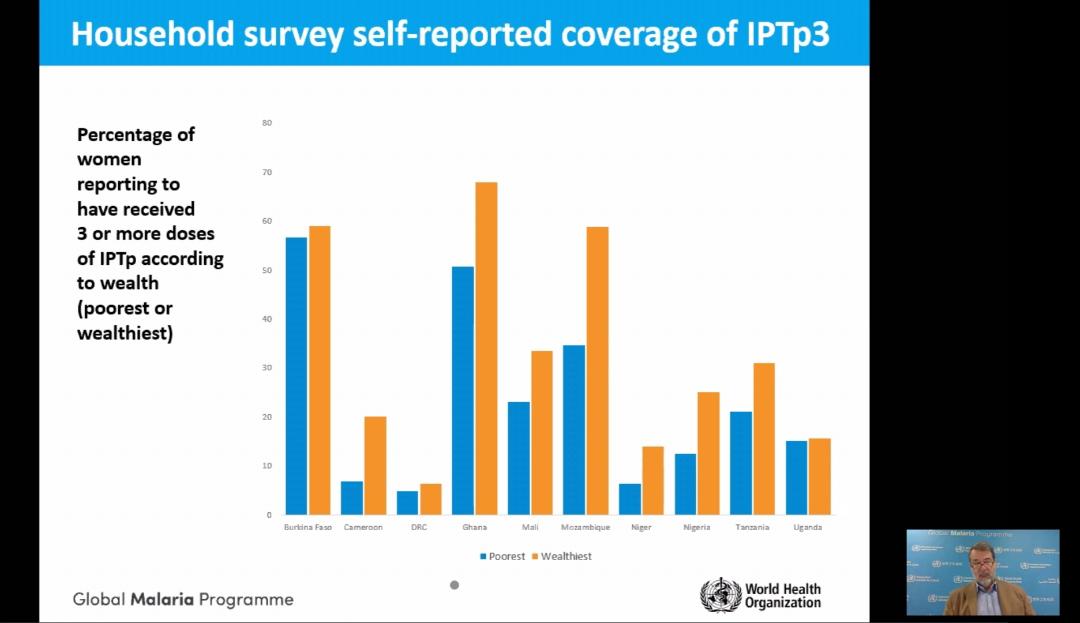
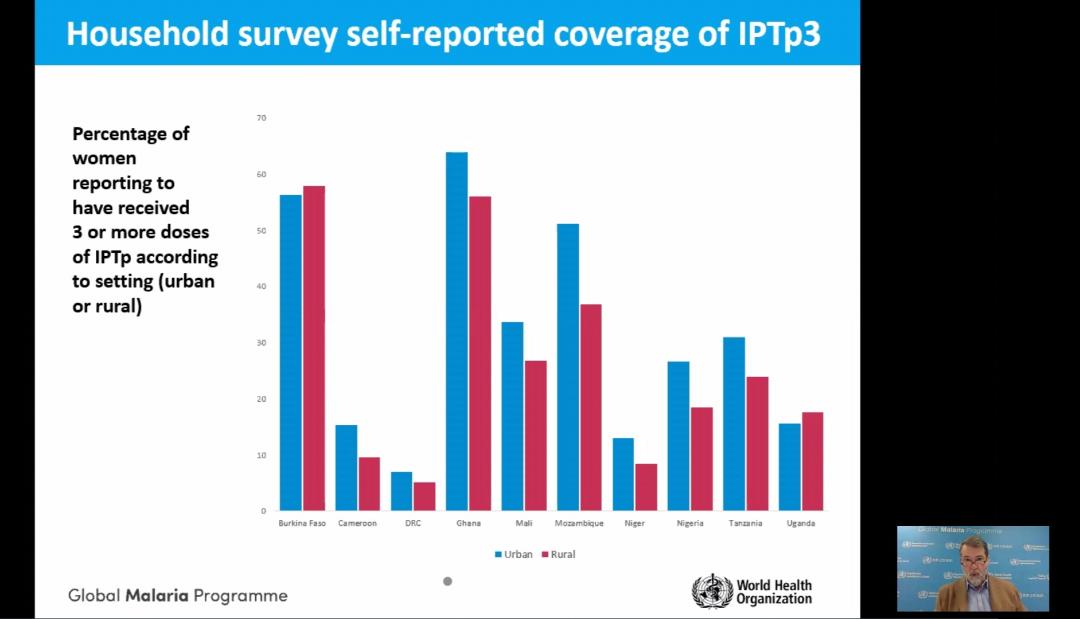
The presentation by Dr Alonso showed the progress in achieving IPTp coverage goals. He also pointed out the social and economic factors that affect access and equity to intermittent preventive treatment of malaria in pregnancy and protection of maternal health (see slides).
Experts say Africa must scale-up malaria protection for pregnant women
 In support of the RBM/AMMREN briefing described above, Sola Ogundipe reports that amidst the COVID-19 pandemic in sub-Saharan Africa, malaria – one of the world’s oldest diseases – is impacting disproportionately on pregnant women and children aged under five.
In support of the RBM/AMMREN briefing described above, Sola Ogundipe reports that amidst the COVID-19 pandemic in sub-Saharan Africa, malaria – one of the world’s oldest diseases – is impacting disproportionately on pregnant women and children aged under five.
For a pregnant woman, her fetus, and the newborn child malaria infection carries substantial risks. Calling for a speedy scale-up to boost protection against malaria for pregnant women in Africa, the Roll Back Malaria, RBM Partnership to End Malaria Working Group is issuing an urgent appeal to leaders and health policymakers to increase access to Intermittent Preventive Treatment during pregnancy, IPTp, among eligible pregnant women in sub-Saharan Africa.
 Along with stakeholders, the RBM Partnership is pushing for scale-up coverage of three doses of IPTp to reach all eligible women in sub-Saharan Africa by 2025. In 2019, according to the RBM Partnership, an estimated 11 million pregnant women in sub-Saharan Africa, or 29 percent of all pregnancies were infected with malaria.
Along with stakeholders, the RBM Partnership is pushing for scale-up coverage of three doses of IPTp to reach all eligible women in sub-Saharan Africa by 2025. In 2019, according to the RBM Partnership, an estimated 11 million pregnant women in sub-Saharan Africa, or 29 percent of all pregnancies were infected with malaria.
How malaria parasites withstand a fever’s heat
The parasites that cause 200 million cases of malaria each year can withstand feverish temperatures that make their human hosts miserable. Now, a team is beginning to understand how they do it. The researchers have identified a lipid-protein combo that springs into action to gird the parasite’s innards against heat shock.
Understanding how malaria protects its cells against heat and other onslaughts could lead to new ways to fight tough-to-kill strains, researchers say. Findings could lead to ways to maximize our existing antimalarial arsenal.
Global Fund and Chevron – United Against HIV, TB and Malaria
The Global Fund and Chevron Corporation have celebrated a 12-year partnership that served as an example of the private sector’s contribution to the fight against infectious diseases and to building resilient health systems. Chevron, a Global Fund partner since January 2008, has supported Global Fund programs against HIX4 tuberculosis and malaria for a total investment of US$60 million in Angola, Indonesia, Nigeria, the Philippines, South Africa, Thailand and Vietnam.
Chevron’s partnership has contributed to helping more than a million people living with HIV access lifesaving antiretroviral therapy; supported efforts to distribute over one million long-lasting insecticide-treated mosquito nets to families to prevent malaria; helped detect thousands of TB cases; promoted education programs for the young, and helped build stronger health systems.
In Nigeria, Chevron’s $5 million investment from October 2017 to December 201 9 supported the national HIV and TB programs, reaching key and vulnerable populations, as well as interventions aimed at strengthening the health system. Sustainable and resilient systems for health are indispensable in the fight against the epidemics of HIV TB and malaria, as well as the first line of defense against new diseases like COVID-19. “With rising cases of COVID-1 9, the stakes are very high. Deaths from HIV, TB and malaria are likely to increase. Investments by partners like Chevron are fundamental.
Artesunate &Children &Diagnosis &Elimination &Plasmodium/Parasite &Private Sector &Resistance &Surveillance Bill Brieger | 25 Sep 2020
Malaria News Today 2020-09-25: Testing, Surveillance, Elimination
Differentiating malaria parasite species is something science can do today, but in the 1800s the debate was over what actually caused the disease. This ability to test and diagnose gives us an important surveillance tool, the 3Ts. News from Ghana is that malaria deaths are reducing, and Guatemala is focusing on elimination. Read more at the links in each section.
Wyss Researchers Develop Malaria Diagnostic Procedure Capable of Differentiating Malaria Species
Researchers at Harvard’s Wyss Institute have developed a new malaria diagnostic test that efficiently detects and can distinguish between species of malaria parasites,
the new procedure significantly improves upon current diagnostic methods, which fail to differentiate between types of malaria. Procedures that only identify Plasmodium falciparum can lead to severe consequences for patients, since other types of malaria are resilient to therapies designed to treat Plasmodium falciparum because they enter a dormant stage in the human liver.
These genes help explain how malaria parasites survive treatment with common drug.
The essential malaria drug artemisinin acts like a “ticking time bomb” in parasite cells—but in the half a century since the drug was introduced, malaria-causing parasites have slowly grown less and less susceptible to the treatment, threatening attempts at global control over the disease.
In a paper published September 23 in Nature Communications, Whitehead Institute Member Sebastian Lourido and colleagues use genome screening techniques in the related parasite Toxoplasma gondii (T. gondii) to identify genes that affect the parasites’ susceptibility to artemisinin. Two genes stood out in the screen: one that makes the drug more lethal, and another that helps the parasite survive the treatment…
Test, Treat, and Track: Strengthening Malaria Response Capabilities in Sierra Leone
A public-private partnership in Sierra Leone aimed to build capacity for testing, treating, and tracking malaria. The post notes the “public-private partnership in Sierra Leone was a collaboration between USAID’s Human Resources for Health in 2030 (HRH2030) program; the U.S. President’s Malaria Initiative (PMI), the Global Fund to Fight AIDS, Tuberculosis and Malaria; the Pharmacy Consultancy of Sierra Leone; and the Sierra Leone National Malaria Control Program”.
NMCP partnered with pharmacies to strengthen their malaria case management capabilities. Anitta and her colleague Brenda Stafford, a trained pharmacist and Procurement and Supply Management officer, led the initiative, going pharmacy-to-pharmacy to train staff on the NMCP’s “Three T” approach: Test, Treat, and Track.
To address the first T, private pharmacies were given free malaria rapid diagnostic tests. According to 2016 data, only half of children under-five with fever received appropriate malaria testing. For the second T, the pharmacists were trained on malaria prevention, treating patients with uncomplicated malaria, and referring patients with severe malaria to health facilities. As for the last T, tracking malaria test results is key in forecasting the spread of disease. NMCP provided pharmacies with two forms that track results: a registry form to track patient information and a summary form which aggregates that into monthly data reports
Miasma War over cause of malaria was heated
Nicole Layton of the Chowan Herald reported that in the 1850s, two North Carolina doctors had a heated and protracted battle over the cause of malaria in the state. This Miasma War is so famous. So what the heck is Miasma? Those fans of Charles Dickens can tell you that at one point miasmas were thought to be the main vector for disease transmission.
During Dickens’ time in London the air was very foul and rather visible because of wood and coal and it smelled bad due to the lack of indoor plumbing. It was thought that this foul air was the cause of disease. Because our part of North Carolina had a lot of rotting vegetation and swamps there was certainly foul air about and very noticeable illness. So the general thought was that malaria was a result of this bad air.
Escuintla, Guatemala: Clinton Health Access Initiative (CHAI)
CHAI’s Guatemala Malaria Team is supporting the Ministry of Health’s National Malaria Program by providing technical assistance and supporting programmatic planning, execution, monitoring and evaluation of effective interventions in the department of Escuintla in southern Guatemala. Over the past five years, CHAI has engaged in the country and helped them make meaningful steps towards orienting their national strategic plans and systems towards the historical goal of malaria elimination.
CHAI’s Guatemala team works in close coordination with the Ministry of Health and other international partners to design, plan, execute and evaluate the impact of the country´s elimination-focused interventions in Escuintla.
Ghana: Malaria deaths in children under five reduce
Ghana has recorded a reduction in malaria deaths in children under five by 83 per cent over the last eight years, the National Malaria Control Programme (NMCP) has said. The rate of malaria-related deaths reduced from 0.6 per cent in 2012 to 0.1 per cent in 2019, showing significant inroads in malaria-related deaths among children. Malaria-related deaths of all ages also reduced by 2,799 in 2012 to 333 by end of 2019, representing an 88 per cent reduction.
Cholera &commodities &Community &coronavirus &Costs &COVID-19 &Culture &Epidemiology &Guidelines &Health Systems &HIV &Microscopy &Mosquitoes &Plasmodium/Parasite &Refugee &Sahel &Seasonal Malaria Chemoprevention &Surveillance &Tuberculosis Bill Brieger | 22 Sep 2020
Malaria News Today 2020-09-22: covering three continents
Today’s stories cover three continents including Surveillance for imported malaria in Sri Lanka, community perceptions in Colombia and Annual Fluctuations in Malaria Transmission Intensity in 5 sub-Saharan countries. In addition there is an overview of microscopy standards and an Integrated Macroeconomic Epidemiological Demographic Model to aid in planning malaria elimination. We also see how COVID-19 is disturbing Seasonal Malaria Chemoprevention activities in Burkina Faso. Read more by following the links in the sections below.
Will More of the Same Achieve Malaria Elimination?
… Results from an Integrated Macroeconomic Epidemiological Demographic Model. Historic levels of funding have reduced the global burden of malaria in recent years. Questions remain, however, as to whether scaling up interventions, in parallel with economic growth, has made malaria elimination more likely today than previously. The consequences of “trying but failing” to eliminate malaria are also uncertain. Reduced malaria exposure decreases the acquisition of semi-immunity during childhood, a necessary phase of the immunological transition that occurs on the pathway to malaria elimination. During this transitional period, the risk of malaria resurgence increases as proportionately more individuals across all age-groups are less able to manage infections by immune response alone. We developed a robust model that integrates the effects of malaria transmission, demography, and macroeconomics in the context of Plasmodium falciparum malaria within a hyperendemic environment.
The authors analyzed the potential for existing interventions, alongside economic development, to achieve malaria elimination. Simulation results indicate that a 2% increase in future economic growth will increase the US$5.1 billion cumulative economic burden of malaria in Ghana to US$7.2 billion, although increasing regional insecticide-treated net coverage rates by 25% will lower malaria reproduction numbers by just 9%, reduce population-wide morbidity by ?0.1%, and reduce prevalence from 54% to 46% by 2034. As scaling up current malaria control tools, combined with economic growth, will be insufficient to interrupt malaria transmission in Ghana, high levels of malaria control should be maintained and investment in research and development should be increased to maintain the gains of the past decade and to minimize the risk of resurgence, as transmission drops. © The American Society of Tropical Medicine and Hygiene [open-access]
Microscopy standards to harmonise methods for malaria clinical research studies
 Research Malaria Microscopy Standards (ReMMS) applicable to malaria clinical research studies have been published in Malaria Journal. The paper describes the rationale for proposed standards to prepare, stain and examine blood films for malaria parasites. The standards complement the methods manual(link is external) previously published by the World Health Organization and UNICEF/UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR). The standards aim to promote consistency and comparability of data from microscopy performed for malaria research and hence to strengthen evidence for improvements in malaria prevention, diagnostics and treatment.
Research Malaria Microscopy Standards (ReMMS) applicable to malaria clinical research studies have been published in Malaria Journal. The paper describes the rationale for proposed standards to prepare, stain and examine blood films for malaria parasites. The standards complement the methods manual(link is external) previously published by the World Health Organization and UNICEF/UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (TDR). The standards aim to promote consistency and comparability of data from microscopy performed for malaria research and hence to strengthen evidence for improvements in malaria prevention, diagnostics and treatment.
Microscopy is important in both malaria diagnosis and research. It is used to differentiate between Plasmodium species and stages and to estimate parasite density in the blood – an important determinant of the severity of disease. It is also used to monitor the effectiveness of drugs based on the rate at which parasites recrudesce or are cleared from the blood.
While rapid diagnostic tests have replaced microscopy in some contexts, microscopy remains an essential tool to support clinical diagnosis and research. The standardisation of methods allows direct comparisons from studies conducted across different points in time and location. This facilitates individual participant data meta-analyses, recognised as the gold standard approach to generate evidence for improvements in interventions and hence patient outcomes.
Estimating Annual Fluctuations in Malaria Transmission Intensity and in the Use of Malaria Control Interventions in Five Sub-Saharan African Countries
RTS,S/AS01E malaria vaccine safety, effectiveness, and impact will be assessed in pre- and post-vaccine introduction studies, comparing the occurrence of malaria cases and adverse events in vaccinated versus unvaccinated children. Because those comparisons may be confounded by potential year-to-year fluctuations in malaria transmission intensity and malaria control intervention usage, the latter should be carefully monitored to adequately adjust the analyses. This observational cross-sectional study is assessing Plasmodium falciparum parasite prevalence (PfPR) and malaria control intervention usage over nine annual surveys performed at peak parasite transmission. Plasmodium falciparum parasite prevalence was measured by microscopy and nucleic acid amplification test (quantitative PCR) in parallel in all participants, and defined as the proportion of infected participants among participants tested. Results of surveys 1 (S1) and 2 (S2), conducted in five sub-Saharan African countries, including some participating in the Malaria Vaccine Implementation Programme (MVIP), are reported herein; 4,208 and 4,199 children were, respectively, included in the analyses.
Plasmodium falciparum parasite prevalence estimated using microscopy varied between study sites in both surveys, with the lowest prevalence in Senegalese sites and the highest in Burkina Faso. In sites located in the MVIP areas (Kintampo and Kombewa), PfPR in children aged 6 months to 4 years ranged from 24.8% to 27.3%, depending on the study site and the survey. Overall, 89.5% and 86.4% of children used a bednet in S1 and S2, of whom 68.7% and 77.9% used impregnated bednets. No major difference was observed between the two surveys in terms of PfPR or use of malaria control interventions. © The American Society of Tropical Medicine and Hygiene [open-access]
Community perception of malaria in a vulnerable municipality in the Colombian Pacific
Malaria primarily affects populations living in poor socioeconomic conditions, with limited access to basic services, deteriorating environmental conditions, and barriers to accessing health services. Control programmes are designed without participation from the communities involved, ignoring local knowledge and sociopolitical and cultural dynamics surrounding their main health problems, which implies imposing decontextualized control measures that reduce coverage and the impact of interventions. The objective of this study was to determine the community perception of malaria in the municipality of Olaya Herrera in the Colombian Pacific.
A 41-question survey on knowledge, attitudes, and practices (KAP) related to malaria, the perception of actions by the Department of Health, and access to the health services network was conducted. In spite of the knowledge about malaria and the efforts of the Department of Health to prevent it, the community actions do not seem to be consistent with this knowledge, as the number of cases of malaria is still high in the area.
Use of a Plasmodium vivax genetic barcode for genomic surveillance and parasite tracking in Sri Lanka
Sri Lanka was certified as a malaria-free nation in 2016; however, imported malaria cases continue to be reported. Evidence-based information on the genetic structure/diversity of the parasite populations is useful to understand the population history, assess the trends in transmission patterns, as well as to predict threatening phenotypes that may be introduced and spread in parasite populations disrupting elimination programmes. This study used a previously developed Plasmodium vivax single nucleotide polymorphism (SNP) barcode to evaluate the population dynamics of P. vivax parasite isolates from Sri Lanka and to assess the ability of the SNP barcode for tracking the parasites to its origin.
A total of 51 P. vivax samples collected during 2005–2011, mainly from three provinces of the country, were genotyped for 40 previously identified P. vivax SNPs using a high-resolution melting (HRM), single-nucleotide barcode method. The proportion of multi-clone infections was significantly higher in isolates collected during an infection outbreak in year 2007. Plasmodium vivax parasite isolates collected during a disease outbreak in year 2007 were more genetically diverse compared to those collected from other years. In-silico analysis using the 40 SNP barcode is a useful tool to track the origin of an isolate of uncertain origin, especially to differentiate indigenous from imported cases. However, an extended barcode with more SNPs may be needed to distinguish highly clonal populations within the country.
Coronavirus rumours and regulations mar Burkina Faso’s malaria fight
 By Sam Mednick, Thomson Reuters Foundation: MOAGA, Burkina Faso – Health worker Estelle Sanon would hold the 18-month-old and administer the SMC dose herself, but because of coronavirus she has to keep a distance from her patients. “If I am standing and watching the mother do it, it’s as if I’m not doing my work,” said Sanon, a community health volunteer assisting in a seasonal campaign to protect children in the West African country from the deadly mosquito-borne disease.
By Sam Mednick, Thomson Reuters Foundation: MOAGA, Burkina Faso – Health worker Estelle Sanon would hold the 18-month-old and administer the SMC dose herself, but because of coronavirus she has to keep a distance from her patients. “If I am standing and watching the mother do it, it’s as if I’m not doing my work,” said Sanon, a community health volunteer assisting in a seasonal campaign to protect children in the West African country from the deadly mosquito-borne disease.
Burkina Faso is one of the 10 worst malaria-affected nations in the world, accounting for 3% of the estimated 405,000 malaria deaths globally in 2018, according to the World Health Organization (WHO). More than two-thirds of victims are children under five. Now there are fears malaria cases could rise in Burkina Faso as restrictions due to coronavirus slow down a mass treatment campaign and rumours over the virus causing parents to hide their children, according to health workers and aid officials.
“COVID-19 has the potential to worsen Burkina Faso’s malaria burden,” said Donald Brooks, head of the U.S. aid group Initiative: Eau, who has worked on several public health campaigns in the country. “If preventative campaigns can’t be thoroughly carried out and if people are too scared to come to health centres … it could certainly increase the number of severe cases and the risk of poor outcomes.”
During peak malaria season, from July to November, community health workers deploy across Burkina Faso to treat children with seasonal malaria chemoprevention (SMC). This is the second year the campaign will cover the whole country with more than 50,000 volunteers going door-to-door, said Gauthier Tougri, coordinator for the country’s anti-malaria programme. Logistics were already challenging. Violence linked to jihadists and local militias has forced more than one million people to flee their homes, shuttered health clinics and made large swathes of land inaccessible. Now the coronavirus has made the task even harder, health workers said.
People in Cape Verde evolved better malaria resistance in 550 years
Yes, we are still evolving. And one of the strongest examples of recent evolution in people has been found on the Cape Verde islands in the Atlantic, where a gene variant conferring a form of malaria resistance has become more common.
Portuguese voyagers settled the uninhabited islands in 1462, bringing slaves from Africa with them. Most of the archipelago’s half a million inhabitants are descended from these peoples. Most people of West African origin have a variant in a gene called DARC that protects.
Deadly malaria and cholera outbreaks grow amongst refugees as COVID pandemic strains health systems.
Apart from the strain on health facilities during the pandemic, in some countries such as Somalia, Kenya and Sierra Leone, we are seeing that a fear of exposure to COVID-19 has prevented parents from taking their children to hospital, delaying diagnosis and treatment of malaria and increasing preventable deaths. COVID restrictions in some countries have also meant pregnant women have missed antimalarial drugs. Untreated malaria in pregnant women can increase the risk of anaemia, premature births, low birth weight and infant death. According to the World Health Organization (WHO), 80% of programs designed to fight HIV, tuberculosis and malaria have been disrupted due to the pandemic and 46% of 68 countries report experiencing disruptions in the treatment and diagnosis of malaria.
Indoor Residual Spraying &Integrated Vector Management &ITNs &Monkeys &Mosquitoes &Nigeria &Plasmodium/Parasite &Seasonal Malaria Chemoprevention &Urban &Vector Control Bill Brieger | 21 Sep 2020
Malaria News Today 2020-09-21: Vectors, Cities and Chimpanzees
First we look at how disease can influence urban planning. We have four news stories focus on field activities for vector control from Hyderabad, India, Borno State, Nigeria, Papua New Guinea and CHAD. Finally there is an ancestry article of sorts examining plasmodia in chimpanzees and humans. Click on the links to read full details.
Can Covid-19 inspire a new way of planning African cities?
 Health crises are not new in Africa. The continent has grappled with infectious diseases on all levels, from local (such as malaria) to regional (Ebola) to global (Covid-19). The region has often carried a disproportionately high burden of global infectious outbreaks.
Health crises are not new in Africa. The continent has grappled with infectious diseases on all levels, from local (such as malaria) to regional (Ebola) to global (Covid-19). The region has often carried a disproportionately high burden of global infectious outbreaks.
How cities are planned is critical for managing infectious diseases. Historically, many urban planning innovations emerged in response to health crises. The global cholera epidemic in the 1800s led to improved urban sanitation systems. Respiratory infections in overcrowded slums in Europe inspired modern housing regulations during the industrial era.
Urban planning in Africa during colonisation followed a similar pattern. In Anglophone Africa, cholera and bubonic plague outbreaks in Nairobi (Kenya) and Lagos (Nigeria) led to new urban planning strategies. These included slum clearance and urban infrastructure upgrades. Urban planning in French colonial Africa similarly focused on health and hygiene issues, but also safety and security.
Unfortunately regional experiences with cholera, malaria and even Ebola in African cities provide little evidence that they have triggered a new urban planning ethic that prioritises infectious outbreaks. Our recent research paper discusses three areas that can transform urban planning in the continent to prepare for future infectious outbreaks, using lessons from Covid-19.
The Coronavirus and other viruses like Ebola have always been ‘out there’ in nature.
 But it’s only when we disrupt the natural habitats of the wild animals. Deadly viruses stay beneath the surface and need just one moment of triggering to emerge in the atmosphere and take the world by storm – historian Dr Mark Honigsbaum. The point is we cannot prevent all spillover events or predict precisely when or where the next one will happen. What we can do — and should do often — is invest in local laboratories and diagnostic services so that we can spot unusual outbreaks early and close them down quickly
But it’s only when we disrupt the natural habitats of the wild animals. Deadly viruses stay beneath the surface and need just one moment of triggering to emerge in the atmosphere and take the world by storm – historian Dr Mark Honigsbaum. The point is we cannot prevent all spillover events or predict precisely when or where the next one will happen. What we can do — and should do often — is invest in local laboratories and diagnostic services so that we can spot unusual outbreaks early and close them down quickly
We should note that Plasmidium Knowlesi is an example of a form of malaria from monkeys that arose because of urban expansion on forest habitats.
Hyderabad: People sensitised on mosquito breeding
As part of a novel initiative, every Sunday 10 am, 10 minute programme, the entomology wing of Greater Hyderabad Municipal Corporation conducted awareness drive on mosquito breeding grounds at various places on Sunday. They explained the importance of cleanliness and the ways the mosquito breeding takes place in stagnated water. Speaking on the occasion, Banjara Hills Corporator Gadwal Vijayalaxmi called upon everyone not to allow accumulation of water in containers, utensils and surroundings.
Borno, WHO Administer Malaria Prevention Drug on 2.1m Children
WHO National Coordinator Malaria Emergencies in Nigeria, Dr. Iniabasi Nglas gave the figure during a four round Malaria Chemoprevention Campaigns (MPCs aka SMC) in 25 of the 27 local government areas of Borno State. During the advocacy, Nglas said the IDP camps “are given special attention for there is high threat of malaria infection due to the environment. Record has shown that the treatment has reduced malaria morbidity in the state.” She revealed that during the first cycle, 1.9 million children were targeted but due to high reception 2.1 million children were administered with the drug.
Rotary Against Malaria Distributes Nets in PNG
ROTARY Against Malaria has finally completed its distribution of bed net mosquito nets throughout the Eastern Highlands Province (EHP) after three months. Team leader of Rotary Against Malaria in the province, Helmut Magino, during a ceremony in Goroka, acknowledged his working staff, the Eastern Highlands Provincial Health Authority, district health officers, logistic company Mapai Transport, Summer Institute of Linguistics (SIL)
and the communities in Papua New Guinea.
“Without these partners, our work in distributing mosquito nets wouldn’t have been successful,” Mr Magino said. “Mapai Transport assisted with vehicles to travel to the remote parts in Okapa, Henganofi and Lufa. “SIL assisted with distribution via airplane to remote parts which are not connected by road like in Obura-Wonenara district.” The volunteer-run organisation funded by Global Fund, a US-based organisation, distributed 145,900 mosquito nets in the province. “We distributed around 45,000 nets to Okapa and Lufa, 35,000 to Obura-Wonenara and 66,900 to rural areas in Goroka district. “We will visit EHP again next year to distribute nets …”
Donating Emergency IRS Supplies to CHAD
Last week, despite the COVID-19 pandemic, a Hercules military transport aircraft took off from an Israeli military base in the south, filled to capacity with items donated by Israeli Flying Aid IFA and the American Jewish Committee (AJC) — 2,000 six-person tents, personal protection equipment (PPE) for medical teams, backpack sprayers to eradicate malaria-carrying mosquitos, and more.
Why humans can run marathons and apes cannot (implication for plasmodium species)
Chimpanzees share more than 99 percent of their genes with modern humans, but the CMAH gene is one of the areas of difference. Two to three million years ago, gorillas, chimpanzees, and other primates were dying from a type of malaria called Plasmodium reichenowi (Science, 2011;331:540-542). At that time, all primates had a surface protein called Neu5Gc on their cells that was made from Neu5Ac. Then along came a primate with a gene that had lost its ability to make Neu5Gc from Neu5Ac, so it had only Neu5Ac (Proc Natl Acad Sci USA, Sept 6, 2005;102(36):12819-12824).
That pre-human did not die from malaria like other primates, his and her children lived and proliferated, and today their descendants (all humans) have a gene that makes Neu5Ac instead of Neu5Gc. As often happens in nature, the malaria parasite then modified its genetic makeup into a variant called Plasmodium falciparum which can infect humans, but not chimpanzees, so today humans can be infected only with Plasmodium falciparum and chimpanzees can be infected only with Plasmodium reichenowi. This same genetic mutation gave homo sapiens greater endurance so they were able to run long distances while the apes could not, which gave humans an advantage in hunting for food (J Hum Evol, 2014;66:64-82).
COVID-19 &Drug Development &Elimination &Funding &Plasmodium/Parasite &Resistance &Vaccine Bill Brieger | 17 Sep 2020
Malaria News Today 2020-09-17
 Leading off our news update are findings from eastern Kenya about a genetic blood grouping that may help prevent malaria. While government leaders in the Asia-Pacific region committed to eliminating malaria, a report from the Gates Foundation explains how COVID-19 has set progress back and without new tools it may take more than 25 years to rid the world of the disease. In that light, Novartis is staking its finances on being able to make medicines that will be accessible and fight malaria. More details and the Gates video can be seen by clicking the links below.
Leading off our news update are findings from eastern Kenya about a genetic blood grouping that may help prevent malaria. While government leaders in the Asia-Pacific region committed to eliminating malaria, a report from the Gates Foundation explains how COVID-19 has set progress back and without new tools it may take more than 25 years to rid the world of the disease. In that light, Novartis is staking its finances on being able to make medicines that will be accessible and fight malaria. More details and the Gates video can be seen by clicking the links below.
How Dantu Blood Group protects against malaria
The secret of how the Dantu genetic blood variant helps to protect against malaria has been revealed for the first time by scientists at the Wellcome Sanger Institute, the University of Cambridge and the KEMRI-Wellcome Trust Research Programme, Kenya. The team found that red blood cells in people with the rare Dantu blood variant have a higher surface tension that prevents them from being invaded by the world’s deadliest malaria parasite, Plasmodium falciparum. The findings were published in Nature and could be significant in the wider battle against malaria.
In 2017, researchers discovered that the rare Dantu blood variant, which is found regularly only in parts of East Africa, provides some degree of protection against severe malaria. The intention behind this new study was to explain why. Red blood cell samples were collected from 42 healthy children in Kilifi, Kenya, who had either one, two or zero copies of the Dantu gene. The Dantu variant created cells with a higher surface tension—like a drum with a tighter skin. At a certain tension, malaria parasites were no longer able to enter the cell.
Novartis ties bond sale to malaria treatment access in sustainability push
Novartis raised 1.85 billion euros on Wednesday from the sale of a bond on which interest payments will rise if the drugmaker fails to expand access to medicines and programmes to combat malaria and leprosy in a number of developing countries.
Investors are increasingly pushing companies to improve their track record on environmental, social and governance (ESG) issues while sustainable investing grows in popularity, spurring an increase in sustainable debt issuance year after year. Novartis’ bond is only the third issue to date to link payments to creditors to company-wide sustainable development targets. By Yoruk Bahceli
Commitment to make Asia Pacific a malaria free region
Government officials from across Asia Pacific have come together during virtual Malaria Week 2020, to reaffirm their commitment to eliminating malaria and strengthening health systems to keep the region safe from health threats. Embracing the theme of “Inclusion. Integration. Innovation.”, officials called for increased collaboration and action to accelerate towards the goal of ending malaria in the region by 2030, at a time when major gains and regional progress are under threat due to disruptions caused by Covid-19.
It could take up to 25 years to eradicate malaria from Africa – Bill Gates
Bill Gates said: “Moving to malaria which is a very awful disease not just to the kids it kills but many kids whose brains are permanently damaged, the economic effects you have with malaria. If we don’t have new tools like vaccines or new ways of killing mosquitoes, it would probably take more than 25 years to get rid of malaria. If we get the new tools and they work, we think it can be done in under 20 years. So the malaria field is both trying to keep the number of deaths down, and we have to deal with the resistance that comes up, that the mosquitoes develop.”
Gates Foundation Report notes that, “We’ve been set back about 25 years in about 25 weeks.” Extreme poverty increased 7% because of COVID-19. In a video included in the Foundation’s report, Bill Gates explains how COVID-19 disrupts the fight against malaria.
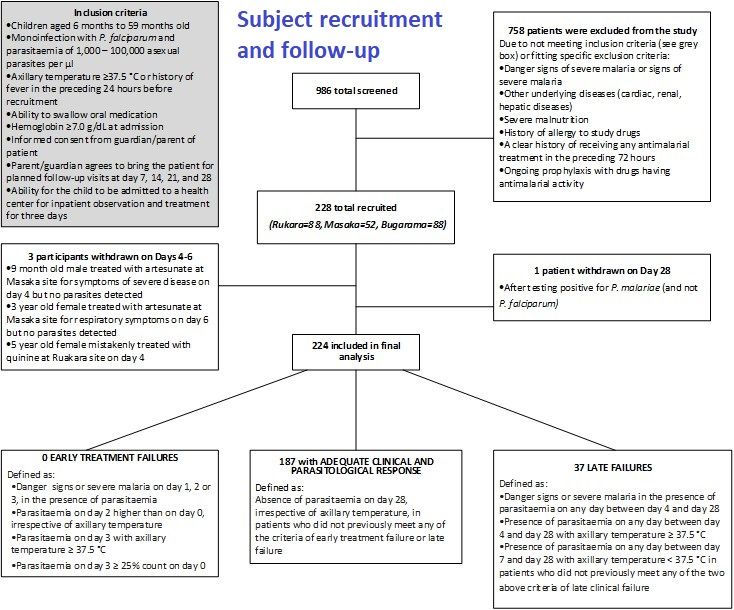
Case Management &Children &Diagnosis &Plasmodium/Parasite Bill Brieger | 23 Nov 2019
Efficacy of artemether-lumefantrine for the treatment of uncomplicated Plasmodium falciparum infection in Rwanda, 2018
 The Efficacy of artemether-lumefantrine for the treatment of uncomplicated Plasmodium falciparum infection in Rwanda, 2018 was investigated by Aline Uwimana, Noella Umulisa, Eric S. Halsey, Meera Venkatesan, Tharcisse Munyaneza, Rafiki Madjid Habimana, Ryan Sandford, Leah Moriarty, Emily Piercefield, Zhiyong Zhou, Samaly Souza, Naomi Lucchi, Daniel Ngamije, Jean-Louis N Mangala, William Brieger, Venkatachalam Udhayakumar, Aimable Mbituyumuremyi.* The results were presented at the 68th Annual Meeting of the American Society of Tropical Medicine and Hygiene and are seen below.
The Efficacy of artemether-lumefantrine for the treatment of uncomplicated Plasmodium falciparum infection in Rwanda, 2018 was investigated by Aline Uwimana, Noella Umulisa, Eric S. Halsey, Meera Venkatesan, Tharcisse Munyaneza, Rafiki Madjid Habimana, Ryan Sandford, Leah Moriarty, Emily Piercefield, Zhiyong Zhou, Samaly Souza, Naomi Lucchi, Daniel Ngamije, Jean-Louis N Mangala, William Brieger, Venkatachalam Udhayakumar, Aimable Mbituyumuremyi.* The results were presented at the 68th Annual Meeting of the American Society of Tropical Medicine and Hygiene and are seen below.
 Background: In Rwanda, there were 4,195,013 confirmed malaria cases and 341 malaria-related deaths in 2018[1]. Regular monitoring of artemisinin-based combination therapy efficacy is important to assess drug efficacy and for timely detection of the emergence of antimalarial drug resistance. In Rwanda, national policy is to routinely monitor the first-line antimalarial per World Health Organization (WHO) guidelines[2] The most recent therapeutic efficacy results in Rwanda showed an efficacy of the first-line antimalarial, artemether-lumefantrine (AL), of >97% in Masaka and Ruhuha in a study conducted from 2013 to 2015[3]
Background: In Rwanda, there were 4,195,013 confirmed malaria cases and 341 malaria-related deaths in 2018[1]. Regular monitoring of artemisinin-based combination therapy efficacy is important to assess drug efficacy and for timely detection of the emergence of antimalarial drug resistance. In Rwanda, national policy is to routinely monitor the first-line antimalarial per World Health Organization (WHO) guidelines[2] The most recent therapeutic efficacy results in Rwanda showed an efficacy of the first-line antimalarial, artemether-lumefantrine (AL), of >97% in Masaka and Ruhuha in a study conducted from 2013 to 2015[3]
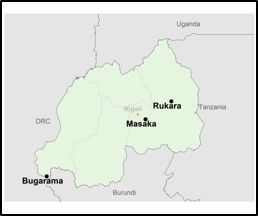
Methods: This was an Efficacy trial based on the standard WHO in vivo protocol[2]. Three sites (see map) were selected in Rwanda. Artemether-lumefantrine (AL) was given twice daily; each dose given under observation for 3 days. Participants were treated with AL and followed for 28 days from March 2018 to December 2018.
 PCR correction, differentiating recrudescence from reinfection in late treatment failure samples, was performed using genotyping of seven neutral microsatellites. Microsatellite data were analyzed using a previously published algorithm that assigns each late treatment failure a posterior probability of recrudescence[4]
PCR correction, differentiating recrudescence from reinfection in late treatment failure samples, was performed using genotyping of seven neutral microsatellites. Microsatellite data were analyzed using a previously published algorithm that assigns each late treatment failure a posterior probability of recrudescence[4]
- Primary Endpoint: 28-day PCR-corrected efficacy
- Secondary Endpoints: 28-day uncorrected efficacy, day 3 parasitemia
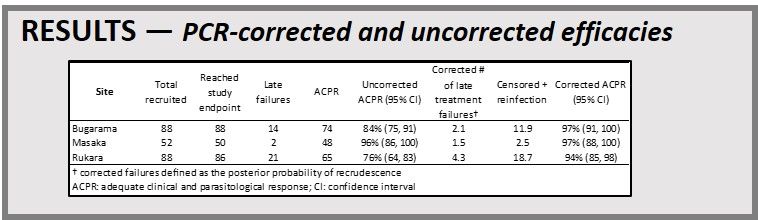 PCR-corrected and uncorrected efficacies are seen to the left. Kaplan Meier Curves are presented. Uncorrected (top) and PCR-corrected (bottom) survival functions for time until failure for a 2018 therapeutic efficacy study using artemether-lumefantrine in three Rwandan study sites; ACPR:
PCR-corrected and uncorrected efficacies are seen to the left. Kaplan Meier Curves are presented. Uncorrected (top) and PCR-corrected (bottom) survival functions for time until failure for a 2018 therapeutic efficacy study using artemether-lumefantrine in three Rwandan study sites; ACPR: 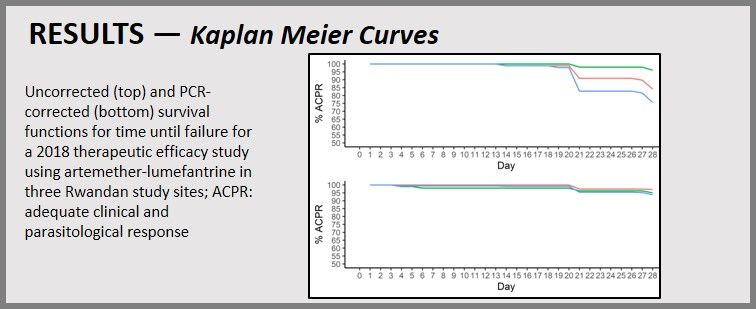 adequate clinical and parasitological response. Day 3 Parasitemia was identified. Two sites, Masaka and Rukara, had > 10% of subjects with parasites detectable on day 3, a WHO criteria for suspected artemisinin resistance.
adequate clinical and parasitological response. Day 3 Parasitemia was identified. Two sites, Masaka and Rukara, had > 10% of subjects with parasites detectable on day 3, a WHO criteria for suspected artemisinin resistance.
 With PCR-corrected efficacies greater than the 90% cut-off recommended by WHO, AL remains an effective antimalarial to treat uncomplicated P. falciparum in Rwanda
With PCR-corrected efficacies greater than the 90% cut-off recommended by WHO, AL remains an effective antimalarial to treat uncomplicated P. falciparum in Rwanda
More than 10% of subjects had day 3 parasitemia at two sites; the relationship with this finding and k13 mutations observed in this study was presented in ASTMH poster LB-5295 (Friday, November 22, 2019).
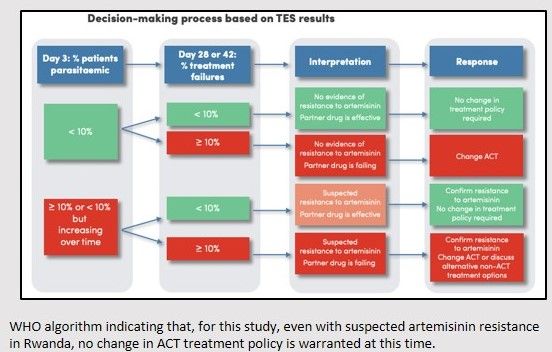 Periodic antimalarial efficacy monitoring in Rwanda should be maintained, and future studies should incorporate additional methods to assess parasite clearance times and presence of molecular markers of resistance. WHO algorithm indicating that, for this study, even with suspected artemisinin resistance in Rwanda, no change in ACT treatment policy is warranted at this time.
Periodic antimalarial efficacy monitoring in Rwanda should be maintained, and future studies should incorporate additional methods to assess parasite clearance times and presence of molecular markers of resistance. WHO algorithm indicating that, for this study, even with suspected artemisinin resistance in Rwanda, no change in ACT treatment policy is warranted at this time.
References
- Rwanda Malaria and Other Parasitic Diseases Division, Rwanda Biomedical Center, HMIS data, 2018.
- WHO, Methods for Surveillance of Antimalarial Drug Efficacy, 2009.
- Uwimana A, Efficacy of artemether–lumefantrine versus dihydroartemisinin–piperaquine for the treatment of uncomplicated malaria among children in Rwanda: an open-label, randomized controlled trial, Trans R Soc Trop Med Hyg; doi:10.1093/trstmh/trz009; 2019.
- Plucinski MM, Morton L, Bushman M, Dimbu PR, Udhayakumar V. Robust algorithm for systematic classification of malaria late treatment failures as recrudescence or reinfection using microsatellite genotyping. Antimicrob Agents Chemother;59:6096–100; 2015.
Contact Information: Aline Uwimana, MD: aline.uwimana@rbc.gov.rw and Eric Halsey, MD: ycw8@cdc.gov
*Affiliations: Malaria and Other Parasitic Diseases Division, Rwanda Biomedical Centre, Kigali, Rwanda; Maternal and Child Survival Program/JHPIEGO, Baltimore MD, USA; The US President’s Malaria Initiative, Atlanta, Georgia, USA; Malaria Branch, Centers for Disease Control and Prevention, Atlanta, Georgia, USA; US President’s Malaria Initiative, Washington DC, USA; National Reference Laboratory, Rwanda Biomedical Centre, Kigali, Rwanda; US Peace Corps, Kigali, Rwanda; US President’s Malaria Initiative, Kigali, Rwanda; WHO Rwanda Office, Malaria and Neglected Tropical Diseases Programs, Kigali, Rwanda; The Johns Hopkins University, Bloomberg School of Public Health, Department of International Health, Baltimore, MD, USA
Epidemiology &History &Migration &Mosquitoes &Plasmodium/Parasite Bill Brieger | 12 Oct 2019
What to Observe on October 12th? Malaria’s Arrival in the Americas
Controversy exists about what historical event should be observed in the USA on 12th October. Ernest Faust explained many years ago that, “there is neither direct nor indirect evidence that the malaria parasites existed on this continent prior to the advent of the European conquerors,” while at the same time in the 16th through 18th Centuries, malaria  was common in England, Spain, France, Portugal and other European nations that arrived in the “New World.” Initially, with the first voyage of Columbus the European explorers and settlers brought the disease, primarily Plasmodium vivax, while the slave trade brought P. falciparum.
was common in England, Spain, France, Portugal and other European nations that arrived in the “New World.” Initially, with the first voyage of Columbus the European explorers and settlers brought the disease, primarily Plasmodium vivax, while the slave trade brought P. falciparum.
National Geographic in its May 2007 issue provided the story “Jamestown, The Real Story.” This article reported that, “Colonists carried the plasmodium parasite to Virginia in their blood. Mosquitoes along the Chesapeake were ‘infected’ by the settlers and spread the parasite to other humans.” Thus malaria became one of many imported diseases that decimated the indigenous population. The spread of P. vivax in Jamestown was not surprising since the settlement was “located on marshy ground where mosquitoes flourished during the summer.”
 Recent research has shown that the “Analysis of genetic material extracted showed that the American P. falciparum parasite is a close cousin of its African counterpart.” This research has documented two genetic groups in Latin America, related to two distinct slave routes run by the Spanish empire in the North, West Indies, Mexico and Colombia and the Portuguese empire to Brazil. Indigenous and remote rural populations of Bolivia, Colombia, Ecuador, Peru, Venezuela and Brazil remain at risk today.
Recent research has shown that the “Analysis of genetic material extracted showed that the American P. falciparum parasite is a close cousin of its African counterpart.” This research has documented two genetic groups in Latin America, related to two distinct slave routes run by the Spanish empire in the North, West Indies, Mexico and Colombia and the Portuguese empire to Brazil. Indigenous and remote rural populations of Bolivia, Colombia, Ecuador, Peru, Venezuela and Brazil remain at risk today.
In the South American continent the native American population might have brought Melanesian strains of P. vivax before the Europeans arrived, but colonizers brought new strains from both Europe and Africa, as well as P. falciparum. Clearly, human migration has played an important role in malaria parasite dissemination through the Americas.
But back to the North American Continent where the USA is observing the historical implications of 12th October, Mark Blackmore reminds us that, “Anthropological and archeological data provide no indication of mosquito-borne diseases among the indigenous people of North America prior to contact with Europeans and Africans beginning in the fifteenth century” (Wing Beats Volume 25 Winter 2015). The spread of malaria by European colonizers is certainly not something to celebrate today.