Plasmodium/Parasite Bill Brieger | 14 Oct 2013 08:36 am
Ghanaian school children harbour antibody responses to antigens on the surface of Plasmodium falciparum gametocyte-infected erythrocytes
Bismarck Dinko of the School of Basic and Biomedical Sciences, University of Health and Allied Sciences, Ho, Ghana and his colleagues Teun Bousema and Colin Sutherland from the Department of Immunology and Infection, London School of Hygiene and Tropical Medicine, London, UK. Presented their research findings at the just concluded Multilateral Initiative for Malaria 6th Pan African Malaria Conference in Durban. Bismark Dinko can be contacted as bismcck@gmail.com, bdinko@uhas.edu.gh for more information.
Malaria transmission-reducing interventions are key to malaria control and possibly elimination.1 Therefore, the development of new tools targeting malaria transmission reduction would mean a major leap forward in malaria control efforts. However, little is known about the immune responses directed at circulating P. falciparum gametocytes in the human host, knowledge of which will be useful in developing transmission reducing interventions targeting gametocytes.
Studies in the Gambia showed P. falciparum gametocytes carry antigens (GSA) which were recognized by malaria patients’ antibodies. These anti-GSA antibodies were found to be associated with lower duration of gametocyte carriage in these patients2,3. Thus, we aimed to determine the presence of anti-GSA antibodies in an asymptomatic population and their relevance to gametocytaemia.
The study was conducted in Ahafo Ano South District, Ashanti Region, Ghana. 274 asymptomatic children aged 6-17yrs were screened by microscopy for malaria, 66% were asymptomatic parasite positive. 155 were treated with DHA-piperaquine upon second visit and enrolled. Enrolled children were followed-up for finger-prick blood donation weekly for 1 month.

Developing stages of P. falciparum gametocytes in culture
Gametocytaemia were determined by Microscopy and QT-NASBA.Gametocytes were produced according to established protocols.3 Mature stage V gametocytes were magnet-purified and tested with plasma samples for antibody recognition by flow cytometry as described elsewhere3 and we present here a summary of the findings.

Prevalence of asexual parasites and gametocytes in malaria asymptomatics
From a cohort of 113 children, all the children harboured plasma antibody responses that recognized GSA on a proportion of mature gametocyte-infected RBCs of 3D7 by flow cytometry. However, 56% of the children exhibited strong antibody responses to GSA (immune response above the median within the cohort per sampling time) by both the proportion of mature gametocytes bound to antibodies and the intensity of the antibody binding to GSA. Longitudinal data provided an additional 10% developing strong GSA responses during the 1 month follow-up.
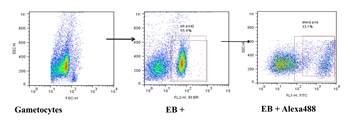
Plasma antibodies recognised mature gametocyte-infected RBCs. Serum from asymptomatic individuals were incubated with mature stage V gametocytes (or trophozoites) and analysed by flow cytometry. Parasites were dual labelled with Alexflour conjugate directly recognizing human IgG and EB staining nuclear DNA. Antibody binding was estimated from the percentage of cells with both EB and Alexaflour, and quadrant settings is based on the single staining controls for EB and Alexaflour.
There were some children with antibody responses fluctuating around the median immune response within the cohort. Children with GSA antibodies present at enrolment were less likely to develop new gametocytaemia at subsequent visits (odds ratio = 0.29, 95% CI 0.06 – 1.05; P = 0.034).
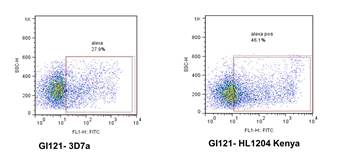
Plasma antibodies from Ghana recognised mature gametocyte-infected RBCs from recent patient isolate from Kenya (HL1204)
3D7 is a laboratory adapted parasite line, so a selection of positive plasma samples was tested against mature gametocyte preparations from HL1204, and strong plasma antibody binding was again shown. No binding to the surface of RBCs infected with immature gametocytes of HL1204 was detected.
In conclusion, a proportion of malaria-infected children carry antibodies that recognized cultured stage V P. falciparum gametocytes from 3D7 and clinical isolates. Strong plasma antibody responses may contribute to gametocytaemia control in vivo. Further work is currently being carried out to identify GSA in collaboration with colleagues at Johns Hopkins School of Public Health, Baltimore, USA.
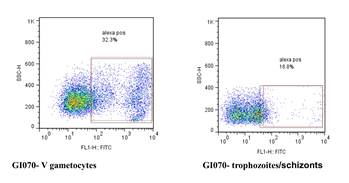
Antibody recognition to the surface of gametocyte-infected RBCs is distinct from the surface of trophozoite-infected RBCs in some children
References
- Alonson et al., 2011. PLoS Med, 8, e1000406.
- Sutherland, 2009, Mol Biochem Parasitol, 166, 93-8.
- Saeed et al., 2008, PLoS One, 3, e2280.
Note that Bismarck Dinko was supported by a MIM travel award.